înapoi la lista
Microdeleții braț lung cromozom Y (AZFa, AZFb, AZFc)
Informaţii generale
Infertilitatea este o problema majora de sanatate care afecteaza 10-15% dintre cupluri, iar in aproximativ jumatate din cazuri poate fi identificat un factor masculin.
Multe cazuri de infertilitate masculina sunt asociate cu oligozoospermie (reducerea productiei de spermatozoizi) sau azoospermie (absenta spermatozoizilor in ejaculat)2. Astfel de alterari in producerea de spermatozoizi pot fi corelate cu modificari histopatologice testiculare de severitate diferita, de la absenta completa a celulelor germinale („sindromul existentei doar a celulelor Sertoli” -SCO) la hipospermatogeneza si oprirea maturatiei. Pe de alta parte, afectarea spermatogenezei se poate datora mai multor cauze: afectiuni sistemice, criptorhidie, tulburari endocrinologice, obstructia/absenta cailor seminale, infectii, medicamente, iradiere etc. Cel putin la un subgrup din aceasta categorie mare de persoane cu defect de spermatogeneza (~ 30%) este implicata o cauza genetica, ceea ce inseamna ca una sau mai multe gene care influenteaza spermatogeneza si-au pierdut functia normala15. Sunt descrise astfel trei cauze genetice de infertilitate masculina:
–anomalii cromozomiale: prezente la 5-10% dintre barbatii cu oligozoospermie si 10-15% dintre cei cu azoospermie; cea mai cunoscuta anomalie este sindromul Klinefelter (47, XXY);
–microdeletii ale cromozomului Y: prezente la 10-18% dintre pacientii cu oligo-sau azoospermie severa;
–mutatiile genei fibrozei chistice (CFTR): se transmit autozomal-recesiv; anumite mutatii pot determina: absenta congenitala bilaterala a canalului deferent (CBAVD), absenta unilaterala a acestuia fara manifestari pulmonare sau pancreatice (CUAVD) sau obstructia canalului deferent; varianta 5T constituie o mutatie comuna asociata cu CBAVD9.
Microdeletiile cromozomului Yconstituie o cauza genetica relativ recent descoperita de infertilitate. Majoritatea acestor anomalii apar de novo si datorita fenotipului asociat cu infertilitate nu sunt de regula transmise descendentilor10.
Detectia microdeletiilor este importanta in special pentru pacientii care intentioneaza sa beneficieze de o reproducere asistata prin ICSI (injectia intracitoplasmatica de sperma) deoarece exista riscul ca aceste anomalii genetice sa fie transmise si descendentilor, ca urmare a suntarii mecanismului fiziologic al fertilizarii2.
Cromozomul Y, unul dintre cei doi gonozomi (cromozomi sexuali), contine 2% din intregul ADN celular si are o lungime de ~60 milioane de perechi de baze. Are rol distinctiv in determinarea sexului masculin prin gena SRY, alcatuita dintr-un singur exon care codifica un factor de transcriptie denumit TDF (testis determining factor)3. Mutatiile survenite la nivelul acestei gene dau nastere unui fenotip feminin cu cariotip 46,XY, iar translocatia genei SRY pe cromozomul X induce un fenotip masculin cu cariotip 46,XX1.
Cei doi cromozomi sexuali X si Y au evoluat dintr-o pereche de cromozomi ancestrali in urma cu cateva sute de milioane de ani; in timp ce cromozomul X a pastrat multe din proprietatile unui autozom (cromozom non-sexual), cromozomul Y si-a pierdut majoritatea genelor, reducandu-si mult dimensiunile (vezi figura 1)5.

Fig.1 Cromozomul X (stanga) si cromozomul Y (dreapta)
Genome biology: Tales of the Y chromosome
Huntington F. Willard, Nature 423, 810-813 (19 June 2003).
Astfel, functia genetica actuala a cromozomului Y este limitata la inducerea dezvoltarii masculine in cursul vietii embrionare si la mentinerea spermatogenezei la adult. La cele doua capete ale cromozomului Y exista regiuni scurte ce sunt identice cu regiunile corespondente de pe cromozomul X, ceea ce indica schimbul de material genetic intre aceste zone (recombinare) in cursul meiozei; cele doua regiuni pseudoautozomale au fost identificate prin bandare cromozomiala si denumite PAR1 si PAR2 (2.6 Mb si, respectiv, 320 kb). Totusi, mai mult de 95% din materialul genetic al cromozomului Y este specific sexului masculin si nu este implicat in procesul de recombinare X-Y – regiune non-recombinanta (non-recombining region NRY). In 2003, Skaletsky si colaboratorii sai au secventiat aceasta regiune si, deoarece au descoperit ca la acest nivel se produce un tip special de recombinare genica, au inlocuit denumirea NRY cu regiunea specifica sexului masculin (male specific region of Y, MSY).
MSY constituie de fapt un mozaic de secvente heterocromatice si eucromatice. Heterocromatina, considerata inerta din punct de vedere genetic, cu un continut inalt de secvente repetitive, este distribuita in principal in portiunea distala a bratului lung al cromozomului Yq ; mai este intalnita in zona centromerica, dar si la nivelul unei portiuni de ~400 kb care intrerupe eucromatina din regiunea proximala Yq. Eucromatina, acea regiune care contine majoritatea genelor (~23 Mb: 8 Mb pe bratul scurt Yp si ~14.5 Mb pe bratul Yq) este alcatuita din 3 clase de secvente nucleotidice:
● regiunea X-transpusa (3.4 Mb derivate prin transpozitie din fostele gene X-linkate);
● regiunea X-degenerata (8.6 Mb derivate din autozomul ancestral);
● secventele ampliconice (10.2 Mb), Y-specifice, caracterizate prin perechi de secvente aproape identice (99.9% similitudine), organizate in palindroame masive (secvente care sunt citite la fel pe ambele catene ale ADN-ului dublu helicoidal); fiecare palindrom prezinta doua brate ce pornesc dintr-un punct central, cu o simetrie in oglinda. Palindroamele MSY, desemnate P1-P8, sunt extrem de mari, cu lungimi ale bratului ce variaza intre 9 Kb (P7) si 1.45 Mb (P1); ca urmare a structurii lor speciale secventele ampliconice sufera un proces de recombinare denumit conversie genica – transfer non-reciproc de material genetic intre secvente duplicate din cadrul aceluiasi cromozom. Deoarece regiunea MSY nu participa la crossing-over-ul X-Y din timpul meiozei, aceasta este lipsita de mecanismul de inlocuire a mutatiilor sau a rearanjarilor structurale cu secvente normale. Se presupune ca fenomenul de conversie genica intre secventele palindromice (recombinarea Y-Y) serveste drept mecanism de refacere a secventelor normale care au devenit nefunctionale pe un brat al palindromului (vezi figura 2).
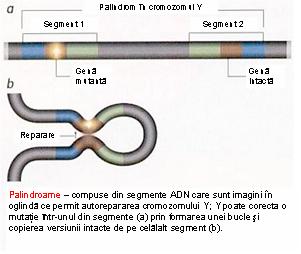
Fig 2. Palindrom in cromozomul Y.
(Adaptare dupa Y Chromosome. http://universe-review.ca/R11-14-YChromosome.htm)16.
Pe langa mentinerea integritatii genelor aceasta structura neobisnuita poate constitui baza structurala pentru deletii sau rearanjari; se crede ca deletiile sunt rezultatul recombinarii omoloage intre secvente repetitive identice cu pierdere de material genetic1;4.
Marea majoritate a regiunilor cromozomului Y (57 din 60 Mb) are o rata redusa de recombinare si se transmite in bloc de la o generatie la alta. Markerii cromozomiali cu o rata mutagena inalta, cum ar fi regiunile microsatelite, sunt variabile in toate populatiile, iar o combinatie specifica de alele poate fi utilizata pentru a defini un haplotip. Prin utilizarea cromatografiei de lichid sub inalta presiune denaturanta (DHPLC) au fost depistate 153 haplotipuri la nivelul cromozomului Y uman. Structura cromozomului Y care se gaseste in GenBank apartine haplotipului R1. Notiunea de haplotip este importanta deoarece persoanele cu diverse haplotipuri Y pot prezenta fenotipuri diferite (de la normal pana la grade variabile de oligozoospermie) atunci cand apar microdeletii7.
O mare parte dintre genele de pe cromozomul Y au fost identificate relativ recent, fiind clasificate in trei grupuri in functie de localizare, numarul de copii si modul lor de exprimare:
a) gene pseudoautozomale, ale caror secvente sunt identice pe cromozomii X si Y (ca de exemplu ASMTL, MIC2, IL9R) si care sunt exprimate, in marea majoritate a cazurilor, in tesuturi diverse;
b) gene localizate in regiunile NRY care sunt omoloage cromozomilor X si Y (USP9Y, DBY, UTY); aceste gene sunt exprimate ubiquitar, desi unele dintre ele prezinta produsi de transcriptie specifici testiculelor;
c) gene Y-specifice (DAZ, CDY, TSPY); aceste gene sunt prezente in copii multiple, fiind larg distribuite pe cromozomul Y sau grupate intr-o regiune mica; sunt exprimate numai la nivelul testiculelor. O exceptie de la aceasta clasificare o constituie gena SRY care determina dezvoltarea testiculelor: desi este Y-specifica, este prezenta intr-o singura copie si are un mod diferit de exprimare ce este limitat la creasta genitala, celulele Sertoli (atat la fat cat si la adult) si celulele germinale2.
Conform publicatiei autorilor L. Tiepolo si O. Zuffardi (1976), exista o zona in regiunea MSY care detine un rol important in spermatogeneza, deletia acestei regiuni fiind asociata cu azoospermia si implicit cu infertilitatea; aceasta zona a fost denumita abreviat AZF (Azoospermia Factor) cu localizare Yq11.2313.
Cu toate acestea, complexitatea genetica a locusului AZF a putut fi evidentiata doar in urma dezvoltarii tehnicilor STS-PCR (Sequence-Tagged Sites PCR) si de hibridizare Southern Blot care au permis detectarea deletiilor interstitiale submicroscopice (invizibile la nivel citogenetic – microdeletii). Studiile de mapare moleculara au complicat ipoteza initiala a locusului unic pentru spermatogeneza de pe Yq sugerand existenta a 3 regiuni ce pot suferi deletii la barbatii infertili; aceste 3 locusuri au fost denumite AZFa, AZFb si AZFc2 (vezi figura 3).

Fig.3: Reprezentare schematica a cromozomului Y (Adaptare dupa www.uhmc.sunysb.edu).
Genele Y din AZFa sunt importante in perioada de embriogeneza si cea pre-pubertara, avand rol in diferentierea spermatogoniilor; genele Y din AZFb si AZFc au rol in maturarea deplina a spermatogoniilor pana la stadiul de spermatozoizi capabili de fertilizare15. Literatura de specialitate mentioneaza faptul ca pacientii carora le lipseste complet regiunea AZFa nu au celule germinale in epiteliul testicular, diagnosticul clinic fiind „sindromul existentei doar a celulelor Sertoli” (Sertoli-Cell-Only-Syndrome SCO)11.
Alaturi de deletiile care apar in una din cele cele 3 regiuni AZFa, AZFb si AZFc, au fost descrise deletii combinate AZFbc, AZFabc, precum si deletii partiale AZFc (denumite AZFc/gr/gr). Frecventa deletiilor, in cazul pacientilor cu azoospermie neobstructiva este diferita pentru AZFa, AZFb, AZFc, AZFbc si AZFabc: 4.9%, 15.8%, 59.6%, 13.6% si, respectiv, <1%. Se presupune ca aceste deletii ar surveni in cursul gametogenezei sau in faza timpurie preimplantare si ar implica un deficit al enzimelor responsabile de repararea ADN-ului normal7.
Genele din regiunea AZFa
Se estimeaza ca intervalul AZFa se intinde pe o lungime de 792 kb si include doua gene functionale larg exprimate: USP9Y (Y-linked Ubiquitin-Specific peptidase 9) si DDX3Y (DEAD Box polypeptyde 3 Y linked), cunoscuta anterior ca DBY. Rolul exact al genelor din regiunea AZFa nu este inca cunoscut, deoarece mutatiile spontane specifice unei singure gene sunt foarte rar intalnite. Deletia completa a regiunii AZFa este relativ rara (frecventa <2% la barbatii cu defect de spermatogeneza) dar bine documentata, fiind intotdeauna asociata cu sindromul SCO3;12.
Gena USP9Y are o lungime de 170 kb, contine cel putin 46 exoni si codifica o proteina ce functioneaza ca o ubiquitin hidrolaza C-terminala. Desi s-a crezut ca deletia genei USP9Y este asociata cu azoospermia si oligospermia, un studiu recent a demonstrat ca USP9Y nu detine un rol esential in producerea spermatozoizilor si fertilitate (ar contribui doar la eficientizarea acestor procese)8.
DDX3Y ar putea reprezenta gena majora a spermatogenezei in aceasta regiune, este mai frecvent implicata in deletii decat USP9Y, prezentand produsi de transcriptie specifici tesutului testicular alaturi de cei ubiquitari. Gena include 17 exoni si codifica o ARN-helicaza ATP dependenta, insa rolul sau in dezvoltarea celulelor germinale maculine nu este complet definit2.
Genele din regiunea AZFb
Regiunea AZFb cuprinde genele EIF1AY (translation-initiation factor 1A, Y isoform) si RBMY (RNA binding motif on Y). Rolul genei EIF1AY in spermatogeneza este pana acum necunoscut, nefiind raportata nici o deletie specifica a acesteia. Deletiile partiale care includ si locusul RBMY, au fost asociate conform studiilor de specialitate cu hipospermatogeneza, iar deletia completa a AZFb este o cauza frecventa de oprire a maturatiei spermatocitelor, ceea ce sugereaza ca familia de gene RBMY are un rol important in spermatogeneza, fiind exprimata numai la nivelul tesutului testicular14.
Genele din regiunea AZFc
AZFc este regiunea de 3.5 Mb a carei deletie este cel mai frecvent intalnita la barbatii cu azoospermie si oligozoospermie; contine mai multe familii de gene implicate in spermatogeneza, candidatul principal in microdeletii fiind DAZ (“deleted in azoospermia”)11;14. DAZ are o lungime de 48 kb, este alcatuita din cel putin 16 exoni si este exprimata specific in tesutul testicular.
Un complex de trei palindroame, cel mai mare avand o lungime de 3 Mb si 99.97% identitate intre bratele sale cuprinde si regiunea AZFc care sufera frecvent deletie. Cu o lungime totala de 4.5 Mb, acesta contine sase familii distincte de unitati repetitive aproape identice (ampliconi). Ampliconii b, g, u, r si t au 4, 3, 3, 4 si, respectiv, 2 secvente repetitive in cadrul haplotipului R1. Aceste secvente reprezinta repetitii directe, repetitii inversate si palindroame. AZFc contine 12 familii de unitati de transcriptie, toate fiind exprimate in tesutul testicular ca BPY2, DAZ, CDY1, CSPG4LY, GOLGA2LY, TTTY3, TTTY4 etc.
Microdeletiile de la nivelul AZFc pot fi grupate in mai multe subtipuri: deletie completa b2/b4 si deletii partiale g1/g2, r1/r2 si r2/r47 (vezi figura 4).
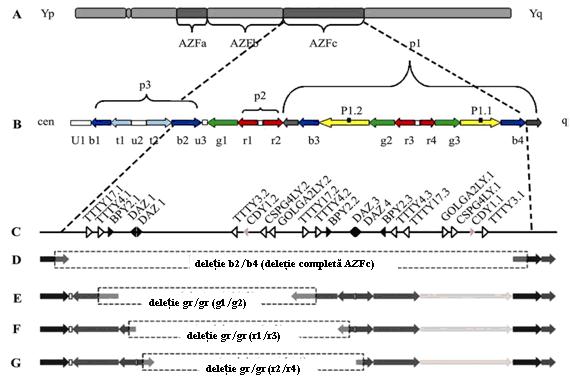
Date recente indica faptul ca deletiile partiale AZFc se produc relativ frecvent; in timp ce unele modificari pot avea relevanta clinica contributia acestora la infertilitatea masculina nu a fost complet analizata12.
Recomandari pentru determinarea microdeletiilor cromozomului Y – evaluarea barbatilor cu azoospermie, oligozoospermie sau infertilitate de cauza neprecizata, pentru stabilirea prognosticului si a optiunilor terapeutice10;12.
Specimen recoltat – sange venos6.
Recipient de recoltare – vacutainer ce contine EDTA ca anticoagulant6.
Cantitate recoltata – 5 mL sange6.
Cauze de respingere a probei – folosirea heparinei ca anticoagulant6.
Stabilitate proba – 7 zile la 2-8ºC6.
Metoda – analiza prin metoda multiplex PCR a regiunilor AZFa, AZFb, AZFc; prin acest test pot fi detectate >95% din deletiile celor 3 regiuni AZF; sunt folositi primeri specifici: sY84 si sY86 pentru regiunea AZFa, sY127 si sY134 pentru regiunea AZFb si SY254 si SY255 pentru AZFc6.
Interpretarea rezultatelor
Microdeletiile Y pot constitui cauza defectului de spermatogeneza. Deoarece fertilitatea este compatibila cu microdeletiile Y (in functie de statusul fertilitatii feminine, fertilizarea naturala se poate produce si in prezenta unui numar redus de spermatozoizi) este mai adecvat sa se considere microdeletiile Y ca fiind cauza oligo-, azoospermiei decat cauza infertilitatii masculine12.
Datele din literatura arata ca deletiile Y relevante sunt asociate de obicei cu un numar de spermatozoizi < 1×106/mL10.
Deletia intregii regiuni AZFa conduce invariabil la sindromul SCO si la imposibilitatea recoltarii de spermatozoizi maturi din epiteliul testicular in vederea injectiei intracitoplasmatice de sperma.
Deletia izolata a genelor USPY9 sau DBY a fost asociata cu fenotipuri testiculare variabile ce nu au fost confirmate de alte grupuri de studiu.
Deletiile complete AZFb si AZFbc se caracterizeaza prin tabloul histologic al SCO sau oprirea maturatiei spermatogoniilor, conditii ce conduc la azoospermie. La fel ca si in cazul deletiei complete AZFa nu se recomanda pacientilor tehnica de reproducere asistata prin ICSI12. Acestia vor fi consiliati de medicul specialist, putand fi luate in considerare optiuni alternative ICSI (de exemplu, donatori de sperma)10.
Deletiile regiunii AZFc (b2/b4) sunt asociate cu un fenotip clinic si histologic variabil si sunt in general compatibile cu spermatogeneza reziduala. Deletiile AZFc pot fi intalnite la barbati cu azoospermie sau oligoazoospermie severa si, in cazuri rare, pot fi transmise descendentilor de sex masculin. De asemenea pacientii cu deletii AZFc pot beneficia de ICSI; copiii lor de sex masculin vor prezenta microdeletii AZFc12.
Limite si interferente
Acest test nu detecteaza toate cauzele de azoospermie sau infertilitate masculina, de aceea un rezultat negativ nu exclude prezenta unui factor genetic sau non-genetic care ar determina tabloul clinic.
Rezultatele testului trebuie interpretate intotdeauna in contextul clinic al pacientului, istoricului familial si al altor date de laborator.
In cazuri rare pot fi detectate polimorfisme care pot conduce la rezultate fals-pozitive sau fals-negative.
Un transplant medular alogenic in antecedente va interfera cu testarea10.
Bibliografie
1. Eberhard Passarge. Genomic Structure of X and Y Chromosomes. In Color Atlas of Genetics, Georg Thieme Verlag KG, 3rd Edition, 2007, 256-257.
2. Foresta C, Moro E,, Ferlin A. Y chromosome Microdeletions and alterations of Spermatogenesis. In Endocrine Reviews, 2001, 22 (2): 226-239.
3. Genetics Home Reference. Chromosome Y. Reference Type: Internet Communication.
4. Helen Skaletskzy et al. The Male-Specific Region of Human Chromosome Y is a mosaic of discrete sequence classes. In Nature 423, 825-836 (19 June 2003).
5. Huntington F. Willard Genome biology: Tales of the Y chromosome. In Nature 423, 810-813 (19 June 2003).
6. Laborator Synevo. Referintele specifice tehnologiei de lucru utilizate. 2010. Ref Type: Catalog.
7. Li Z, Haines C, Han Y. “Micro-deletions” of the human Y chromosome and their relationship with male infertility. In J Genet. Genomics, 2008, 35: 193-199.
8. Luddi A, Margollicci M, Gambera L, Serafini F, Cioni M, De Leo V, Balestri P, Piomboni P. Spermatogenesis in a man with complete deletion of USP9Y. In N Engl J Med. 2009 Feb 26;360(9):881-5.
9. Male Infertility. www.genecare.com. Reference Type: Internet Communication.
10. Mayo Clinic, Mayo Medical Laboratories. Reference Laboratory Services for Health Care Organizations. Y Chromosome Microdeletions, Molecular Detection. www.mayomedicallaboratories.com. 2010. Ref Type: Internet Communication.
11. Navarro-Costa P, Goncalves, Plancha CE. The AZFc region of the Y chromosome: at the crossroads between genetic diversity and male infertility. In Hum. Reprod. Update, 2010, March, 1-18.
12. Simoni M, Bakker E, Krausz C. EAA/EMQN best practice guidelines for molecular diagnosis of y- chromosomal microdeletions. State of the art 2004. In Int J Androl 2004;27: 240-9.
13. Tiepolo L, Zuffardi O. Localisation of factors controlling spermatogenesis in the nonfluorescent portion of the human Y chromosome long arm. In Hum. Genet. 1976, 34:119-124.
14. Vogt HP. Azoospermia factor (AZF) in Yq11: towards a molecular understanding of its function for human male fertility and spermatogenesis. In RMB Online, 2005, 10 (1): 81-93.
15. www.klinikum.uni-heidelberg.de/AZ. Deletionen-und-maennliche-Infertilitaet. Reference Type: Internet Communication.
16.Y Chromosome. http://universe-review.ca/R11-14-YChromosome.htm. Reference Type: Internet Communication.
Vezi tot conținutul
Vezi mai puțin
